研究概要 (内山 英穂)
 動物の発生を研究するには両生類は好適な材料です.卵が大きく遺伝子注入や手術が容易に行えるためです。また、カエルのゲノム解読も今ではかなり進展しました。私はアフリカツメガエルを材料として中胚葉と神経の分化の研究を行っており、特にT-ボックス転写因子群 に注目しています。この因子群はアクチビン、FGF(繊維芽細胞成長因子)やBMP (骨形成タンパク質)といった中胚葉誘導因子により中胚葉誘導がおこると、すぐに発現して細胞分化の引き金をひくMaster Control Genes です。
動物の発生を研究するには両生類は好適な材料です.卵が大きく遺伝子注入や手術が容易に行えるためです。また、カエルのゲノム解読も今ではかなり進展しました。私はアフリカツメガエルを材料として中胚葉と神経の分化の研究を行っており、特にT-ボックス転写因子群 に注目しています。この因子群はアクチビン、FGF(繊維芽細胞成長因子)やBMP (骨形成タンパク質)といった中胚葉誘導因子により中胚葉誘導がおこると、すぐに発現して細胞分化の引き金をひくMaster Control Genes です。
現在研究しているT-box遺伝子 Tbx6 は、正中線では発現せず、正中線から少し脇にそれた部位にあたる沿軸中胚葉および側板中胚様で強く発現しています。単独で胞胚中期の予定外胚葉(アニマルキャップ)に発現させるだけで側板中胚葉様の細胞分化をひき起こしますが、脊索の分泌する背側化因子と同時に発現させると筋肉を分化させます。
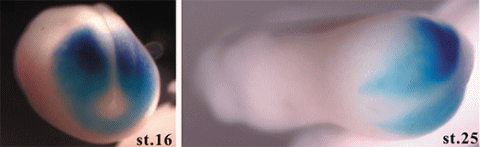
tbx6は、ノックダウン実験により体節や腹側体壁における筋形成と分節形成、さらには頭部軟骨や頭部骨格筋の形成に必須です。Tbx6はpMesogeninというbHLH型転写因子の発現を引き起こし、pMesogeninが体壁筋を分化させます。左の写真はアフリカツメガエルの神経胚と尾芽胚で、濃い青がTbx6rの、また淡い青がTbx6の発現部位です。Tbx6rの発現領域はtbx6の発現領域の中の一部です。カエルではこれら2つの転写因子のパラログが沿軸中胚葉の分化制御を行っています。現在、Crispr/Cas9を用いたノックアウト実験を行っています。
柱のもう1つは、哺乳類の多能性幹細胞 (Pleuripotent stem cell)を用いた細胞分化の研究です。この分野ではマウスの様々なES細胞を材料として、3次元の細胞塊(胚様体)の作成や、液性因子の添加により様々な中胚葉組織を分化させる細かい条件検討を行っています。Tbx6遺伝子を発現させると、神経分化が完全に抑制され、中胚葉分化が昂進することもわかりました。興味のわいた人は是非訪ねてみてください。大学院は他大学出身者も歓迎。
<最近の研究テーマ>
1 T-box型転写因子による細胞分化制御機構
2 SproutyやOrexinの神経分化における役割の解析
3 マウスES細胞用いた種々の中胚葉系、神経系細胞分化条件の確立
| 所属学会 |
日本動物学会、 日本発生生物学会、 日本分子生物学会、 国際発生生物学会 |
| 担当科目・学部 |
自然科学基礎実験C(1年次後期)、再生発生学 (3年次前期)、動物細胞工学(3年次前期)、生命環境専門実習Ⅲ(3年次前期) |
| 担当科目・大学院 |
ゲノムシステム科学概説Ⅱ (博士前期)、 発生システム制御科学Ⅱ (博士前期)、ゲノムシステム科学特別講義Ⅱ(博士後期)、発生システム制御科学講究Ⅱ(博士後期) |
--- Recent Publishing ---
1)塩尻信義ら/共編著 発生生物学 基礎から応用への展開 培風館 2019年4月 (第8章 ボディープランの確立 を執筆)
2) Yano Y, Iimura N, Kojima N, and Uchiyama H. Non-neural and cardiac differentiating properties of Tbx6-expressing mouse embryonic stem cells.(Tbx6を発現するマウス胚性幹細胞のもつ非神経的および心臓分化の性質) Regenerative Therapy, 3, 1-6, 2016.
3) Hasan I, Sugawara S, Fujii Y, Koide Y, Terada D, Iimura N, Fujiwara T, Takahashi KG, Kojima N, Rajia S, Kawsar SMA, Kanaly RA, Uchiyama H, Hosono M, Ogawa Y, Fujita H, Hamako J, Matsui T, and Ozeki Y. MytiLec, a Mussel R-Type Lectin, Interacts with Surface Glycan Gb3 on Burkitt’s Lymphoma Cells to Trigger Apoptosis through Multiple Pathways.(ムラサキイガイのR型レクチンMytiLecはバーキットリンパ腫のGb3表面糖鎖と相互作用して細胞死を引き起こす)Marine Drugs, 13(12), 7377-7389, 2015.
4) Avila FM., Yamaguchi, S., Uchiyama, H., and Micheletto, R. (2011). A λ/30 resolution laser speckle pattern biosensor for dynamic studies on live samples.(生きた標本の動的研究のためのλ/30解像度をもつレーザー斑点模様)Intelligent Sensors, Sensor Networks and Information Processing (ISSNIP) Seventh International Conference, 37-40, 2011.
5) Aihara Y, Hayashi Y, Hirata M, Ariki N, Shibata S, Nagoshi N, Nakanishi
M, Ohnuma K, Warashina M, Michiue T, Uchiyama H, Okano H, Asashima M and Furue MK Induction of neural crest cells from mouse embryonic stem cells in a
serum-free monolayer culture. (無血清単層培養系によるマウス胚性幹細胞から神経堤細胞の誘導)Int. J. Dev. Biol., 54, 1287-1294, 2010.
6) Konno, M.,, Hamazaki, T. S., Fukuda, S., Tokuhara, M., Uchiyama, H., Okazawa, H., Okochi, H., Asashima, M. Efficiently Differentiating Vascular Endothelial Cells from Adipose Tissue Derived Mesenchymal Stem Cells in Serum-Free Culture (脂肪組織に由来する間葉系幹細胞の無血清培養による血管内皮細胞への効率的分化)Biochem. Biophys. Res. Commun., 400, 461-465, 2010.
7) Tazumi, S., Yabe, S., and Uchiyama, H. Paraxial T-box genes, Tbx6 and Tbx1, are required for cranial chondrogenesis and myogenesis.(沿軸中胚葉のT-box遺伝子であるTbx6とTbx1は、頭部筋肉および頭蓋軟骨の発生に必要である)Devl. Biol., 340, 170-180, 2010.
Accession number of X. laevis pcdh18: AB573177
8) Kawsar, S.M.A., R. Matsumoto, Y. Fujii, H. Yasumitsu, H. Uchiyama, M. Hosono, K. Nitta, J. Hamako, T. Matsui, N. Kojima, and Y. Ozeki. Glycan-binding profile and cell adhesion activity of American bullfrog (Rana catesbeiana) oocyte galectin-1. (ウシガエルの卵母細胞 galectin-1 の糖鎖結合プロファイリングと細胞接着活性)Prot. Peptide Lett. 16, 677-684, 2009.
9) Hitachi, K., Danno, H., Tazumi S., Aihara Y., Uchiyama, H., Okabayashi, K., Kondow, A., and Asashima, M. The Xenopus Bowline/Ripply family proteins negatively regulate the transcriptional activity of T-box transcription factors.
(アフリカツメガエルのBowline/Ripplyファミリータンパク質はT-box転写因子の転写活性を抑制的に制御している)Int. J. Dev. Biol. 53, 631-639, 2009.
10) Tazumi, S., S. Yabe, J. Yokoyama, Y. Aihara, and H. Uchiyama. pMesogenin1 and 2 function directly downstream of Xtbx6 in Xenopus somitogenesis and myogenesis.
(ツメガエルの体節形成と筋形成においてpMesogenin1と2はXtbx6の直接の下流で機能している)Dev. Dyn. 237, 3749-3761, 2008.
Dev. Dyn. 238, issue2, page fvii, DD Artpix, 2009 でもとりあげられています
Accession number of pMesogenin2 genomic DNA: AB462327.
Accession number of Xtbx6 exon1-exon2 genomic DNA: AB462326.
11) Hitachi K., Danno H., Kondow A., Ohnuma K., Uchiyama H., Ishiura S., Kurisaki A, and Asashima M. Physical interaction between Tbx6 and mespb is indispensable for the activation of bowline expression during Xenopus somitogenesis.(アフリカツメガエルの体節形成の間、bowlineの活性化にはTbx6とmespbの物理的な相互作用が必要である)Biochem. Biophys. Res. Commun., 372, 607-612, 2008.
12) Hitachi K., Kondow A., Danno H., Inui M., Uchiyama H., and Asashima M. Tbx6, Thylacine1, and E47 synergistically activate bowline expression in Xenopus somitogenesis.(アフリカツメガエルの体節形成においてTbx6、Thylacine1、E47は相乗的にbowlineの発現を活性化する)Devl. Biol., 313, 816-828, 2008.
13) Yabe S., Tazumi S., Yokoyama J., and Uchiyama, H. Xtbx6r, a novel T-box gene expressed in the paraxial mesoderm, has anterior neural-inducing activity.(前方神経誘導活性をもつ、沿軸中胚葉で発現する新規T-box遺伝子Xtbx6r)Int. J. Dev. Biol., 50, 681-689, 2006.
Accession number of Xtbx6r cDNA: AB284536
14) Murakami R., Okumura T., and Uchiyama H. GATA factors as key regulatory molecule in the development of Drosophila endoderm. (ショウジョウバエの内胚葉の発生における鍵調節分子であるGATA因子)Dev. Growth Differ. 47, 581-589, 2005.
15) Hamaguchi T., S. Yabe, H. Uchiyama, and R. Murakami. Drosophila Tbx6-related gene, Dorsocross, mediates high levels of Dpp/Scw signal required for the development of amnioserosa and wing discs. (ショウジョウバエのTbx6に関連した遺伝子Dorsocrossは、amnioserosaと羽成虫原基の発生に必要な高いレベルのDpp/Scwシグナルを仲介する) Devl. Biol. 265, 355-368, 2004.
16) Uchiyama H., T. Kobayashi, A. Yamashita, S. Ohno and S. Yabe. Corrigendum: Cloning and characterization of the T-box gene Tbx6 in Xenopus laevis. Develop Growth Differ. 44, 95-96, 2002.
Uchiyama, H., T.Kobayashi,A.Yamashita,S. Ohno, and S. Yabe. Cloning and characterization of a T-box gene Tbx6 in Xenopus laevis. (アフリカツメガエルにおけるT-box遺伝子Tbx6のクローニングと性格づけ) Develop. Growth Differ. 43, 657-669, 2001.
Accession number of Xtbx6 cDNA: AB091393
17) Uchiyama, H., A. Koda, S. Komazaki, M. Oyama, and S. Kikuyama. Occurrence of immunoreactive activin/inhibin βB in thyrotropes and gonadotropes in the bullfrog pituitary: possible paracrine/autocrine effects of activin B on gonadotropin secretion.
(ウシガエル脳下垂体の甲状腺刺激ホルモン分泌細胞と性腺刺激ホルモン分泌細胞における免疫反応性アクチビン/インヒビンβB鎖の存在: 性腺刺激ホルモン分泌に対するアクチビンBの傍分泌・自己分泌的効果の可能性). Gen. Comp. Endocrinol. 118, 68-76 (2000).
18) Koda A., K. Yamamoto, H. Uchiyama, H. Vaudry, and S. Kikuyama. Effect of activin A and follistatin on the release of pituitary hormones in the bullfrog Rana Catesbeiana.(ウシガエルにおける脳下垂体ホルモン分泌へのアクチビンAの影響) Zool. Sci. 17, 971-975 (2000)
19) Yamamoto T., S. Iemura, S. Takagi, H. Uchiyama, S. Shimasaki, H. Sugino and N. Ueno. Direct binding of follistatin to BMPs inhibits ventral and epidermal cell fates in early Xenopus embryo. (初期アフリカツメガエル胚において骨形成タンパク質とその受容体の複合体へのフォリスタチンの直接結合が,腹側および表皮への細胞運命を抑制する) Proc. Natl. Acad. Sci. USA 95, 9337-9342 (1998).
20) 内山英穂, 浅島 誠. 中胚葉誘導から胚のボディープランへ -胚の頭尾・背腹軸決定のしくみ- 「両生類の発生生物学」 第9章 片桐千明編著、北海道大学出版会 pp187-206 (1998).
研究室先頭ページに戻る